Naoyuki Takeuchi, MD, PhD1, Takeo Tada, MD, PhD2, Masahiko Toshima, MD3 and
Katsunori Ikoma, MD, PhD1
From the 1Department of Rehabilitation Medicine, Hospital of Hokkaido University, Sapporo, 2Department of
Rehabilitation Medicine, Hospital of Sasson, Otaru and 3Department of Rehabilitation Medicine Carres Sapporo,
Hospital of Tokeidai, Sapporo, Japan
OBJECTIVE: The inhibitory role of neuronal networks in motor recovery after stroke remains to be elucidated. We examined the influence of transcallosal inhibition and short intracortical inhibition on motor recovery after stroke. We also investigated the correlation between transcallosal inhibition and mirror activity.
DESIGN: A cross-sectional study.
SUBJECTS: Thirty-eight chronic stroke patients.
METHODS: Transcallosal inhibition was evaluated using single transcranial magnetic stimulation, and short intracortical inhibition was assessed using paired-pulse transcranial magnetic stimulation. Mirror activity was measured during tonic contraction of the contralateral hand.
RESULTS: Transcallosal inhibition from the contralesional to the ipsilesional motor cortex correlated positively with motor function of the paretic hand; in contrast, transcallosal inhibition to the ipsilesional motor cortex correlated negatively with mirror activity of the paretic hand in both cortical and subcortical stroke patients. Short intracortical inhibition of the ipsilesional motor cortex correlated negatively with motor function of the paretic hand in only the subcortical stroke patients.
CONCLUSION: Transcallosal inhibition from the contralesional to the ipsilesional motor cortex may inhibit mirror movements in stroke patients with good motor function. The weak transcallosal inhibition in patients after stroke with poor motor function may be ineffective for inhibiting mirror movement; however, it may have the advantage of facilitating motor recovery.
Key words: stroke; rehabilitation; reorganization; mirror movement; transcallosal inhibition; intracortical inhibition.
J Rehabil Med 2010; 42: 962–966
Correspondence address: Naoyuki Takeuchi, Department of Rehabilitation Medicine, Hospital of Hokkaido University. North 14 West 5 Sapporo 060-8648, Japan. E-mail: naoyuki@med.hokudai.ac.jp
Submitted January 28, 2010; accepted September 1, 2010
Introduction
Stroke alters the neuronal function of the motor cortex adjacent to or distant from the lesion through neuronal networks (1). Transcranial magnetic stimulation (TMS) has been used to detect changes in neuronal function after stroke. Several studies have reported the loss of inhibition in the ipsilesional and the contralesional motor cortex of stroke patients using TMS (2, 3). A decrease in the inhibition contributes to the cortical reorganization by unmasking the latent networks (4); however, whether the disinhibition after stroke is caused by the lesion, whether it reflects a compensatory mechanism, or both, is still poorly understood (1). The change in transcallosal inhibition (TCI) after subcortical stroke has also been assessed using TMS (5). While a recent study has examined the changes in both TCI and intracortical inhibition after stroke (6), it remains unknown whether these neurophysiological parameters are correlated with motor function in both cortical and subcortical stroke and whether the parameters of cortical stroke differ from those of subcortical stroke.
In this study, we evaluated TCI and short intracortical inhibition (SICI) to determine whether these TMS parameters influence motor recovery in both cortical and subcortical stroke. It has been demonstrated previously that although SICI may be reduced in appearance, the inhibitory function may be normal if the excitability function increases (7). Therefore, we measured not only SICI but also short interval cortical excitability (SICE) to evaluate inhibitory and excitatory function in more detail. In addition, we investigated the correlation between TCI from the contralesional to the ipsilesional motor cortex and the mirror activity of the paretic hand. We hypothesized that the change in TCI to the ipsilesional motor cortex after stroke could influence the mirror activity of the paretic hand during non-paretic hand movement.
Methods
The study population comprised 38 first-time chronic stroke patients. Motor function was evaluated using the upper limb subset of the Fugl-Meyer scale (FMS) (8). All the subjects gave written informed consent, and the experimental protocol was approved by the local ethics committee of Hokkaido University Graduate School of Medicine. The patients were classified into the following two subgroups according to brain computed tomography (CT) or MRI findings (Table I): (i) the cortical group, which had stroke lesions involving the sensorimotor cortex or both sensorimotor cortex and subcortical structure; and (ii) the subcortical group, which had lesions located caudal to the corpus callosum, indicating that the corpus callosum was intact.
| Table I. Clinical characteristics |
| | Age, year Mean (SD) | Gender | Paretic side | Duration after stroke, month Mean (SD) | Fugl-Meyer scale, Mean (SD) | *EMG activity of first dorsal interosseous |
| Male n | Female n | Right n | Left n | Non-paretic, μV Mean (SD) | Paretic μV Mean (SD) |
| Cortical group (n = 20) | 61.7 (10.1) | 12 | 8 | 12 | 8 | 46.3 (34.2) | 68.0 (23.4) | 350.8 (210.2) | 155.0 (140.8) |
| Subcortical group (n = 18) | 61.6 (10.3) | 11 | 7 | 10 | 8 | 56.9 (51.9) | 63.9 (21.7) | 395.4 (220.1) | 154.6 (155.5) |
| *Mean rectified EMG activity during maximal tonic contraction. SD: standard deviation; EMG: electromyography. |
TCI was performed using a 70-mm figure-8 coil and Magstim 200 (Magstim Company, Dyfed, UK), and paired-pulse TMS was applied using the same coil and a Bistim device (Magstim Company) that triggered two magnetic stimulators. The coil was placed tangentially over the motor cortex at an optimal site for the first dorsal interosseous (FDI) muscle. The optimal site was defined as the location where stimulation at a slightly suprathreshold intensity elicited the largest motor-evoked potentials (MEPs) in the FDI. The resting motor threshold (rMT) was determined separately for each stimulator and defined as the lowest stimulator output that could activate MEPs with a peak-to-peak amplitude greater than 50 μV in at least half of the 10 trials. We excluded patients for whom MEPs were not detected in the ipsilesional hemisphere from the ipsilesional TMS study section, i.e. patients in whom MEPs were not induced even at 100% stimulator output.
We performed paired-pulse TMS at inter-stimulus intervals (ISIs) of 2, 3, 10 and 15 ms. The intensity of the first conditioning stimulus was 80% rMT and that of the test stimulus was 120% rMT. Ten trials were performed for each ISI and unconditioned trials (controls) were recorded during complete relaxation. The paired stimulation with each ISI was randomly mixed with the control stimulation. The mean peak-to-peak amplitude of the control MEPs and paired MEPs at each ISI was calculated. The mean amplitudes of paired MEPs at ISIs of 2 and 3 ms were averaged to obtain a representative value for SICI and that at ISIs of 10 and 15 ms intervals for intracortical facilitation (ICF). SICI is expressed as the percentage of the degree of inhibition (1 − (paired/control)), and ICF is expressed as the percentage increase (paired/control) in MEPs amplitude. SICE was measured using paired-pulse TMS at an ISI of 2 ms. The intensity of the conditioning stimulus varied between 30% and 80% of MT and was administered randomly at 10% increments; whereas, the intensity of the test stimulus was the same as that for the SICI measurement. MEPs amplitudes at each conditioning stimulus in SICE were expressed as a percentage of the mean amplitude of the control MEPs.
In the TCI procedure, each hemisphere was stimulated 20 times (intensity, 150% rMT) during unilateral maximal tonic contraction of the ipsilateral FDI, while keeping the contralateral upper limb relaxed as described previously (9). Twenty electromyography (EMG) signals of the FDI were rectified and averaged for evaluation of TCI. The mean amplitude of EMG signals prior to the stimulus for 100 ms was defined as the background activity. TCI was quantified by the period of relative EMG suppression after the stimulus, i.e. from the point at which the EMG activity clearly decreased below the background activity to that at which the EMG activity again increased to equal the background activity. The area of suppressed EMG activity was also averaged. TCI was then defined as the percentage of this mean suppressed activity in the background activity. This indicates that the greater the EMG activity suppression, the greater the TCI.
Mirror activity was calculated from the data in the TCI section to avoid the fatigue of stroke patients by additional tests. We rectified and averaged 20 EMG signals of the contralateral FDI muscles (mirror condition) prior to TMS for 100 ms during a maximal tonic contraction of the FDI muscle (active condition). Finally, mirror activity was expressed as a percentage of the mean amplitude of the mirror condition in the mean amplitude of the active condition at the same FDI.
Clinical data were compared between the cortical and subcortical groups by using the Mann-Whitney U test or the χ2 test, depending on the type of variable assessed. For the comparison of TMS parameters, the Kruskal-Wallis test was used. The changes in SICE were evaluated using analysis of variance (ANOVA) for repeated measures, with INTENSITY as a within-subjects factor and STIMULATION SITE as a between-subjects factor. A post-hoc analysis was performed with Bonferroni’s correction. Possible correlations among the various parameters were determined using the Spearman’s correlation test.
Results
There was no significant difference between the cortical and subcortical groups with regard to age, gender, paretic side, duration after stroke, FMS, EMG activity of non-paretic, or EMG activity of paretic (Table I). Table II shows TMS parameters of each hemisphere in the subcortical and cortical groups. We obtained ipsilesional TMS data from 9 patients in the cortical group and 9 patients in the subcortical group. There was no significant difference between the 4 stimulation sites with regard to rMT, amplitude of MEPs, SICI, ICF, or TCI (Table II).
| Table II. Transcranial magnetic stimulation parameters |
| Stimulation site | rMT, % Mean (SD) | Amplitude of MEPs, μV Mean (SD) | SICI, % Mean (SD) | ICF, % Mean (SD) | TCI, % Mean (SD) |
| Ipsilesional hemisphere in cortical group (n = 9) | 52.8 (12.2) | 921.9 (463.6) | 38.4 (50.6) | 169.2 (71.8) | 50.1 (14.0) |
| Ipsilesional hemisphere in subcortical group (n = 9) | 50.9 (9.7) | 556.8 (348.7) | 23.6 (41.7) | 182.6 (160.8) | 53.7 (14.3) |
| Contralesional hemisphere in cortical group (n = 20) | 51.9 (9.1) | 895.0 (451.7) | 25.7 (65.8) | 192.2 (93.6) | 46.2 (15.1) |
| Contralesional hemisphere in subcortical group (n = 18) | 52.9 (8.6) | 813.6 (670.0) | 22.0 (49.6) | 239.6 (139.5) | 58.7 (14.6) |
| rMT: resting motor threshold; MEPs: motor evoked potentials; SICI: short intracortical inhibition; ICF: intracortical facilitation; TCI: transcallosal inhibition; SD: standard deviation. |
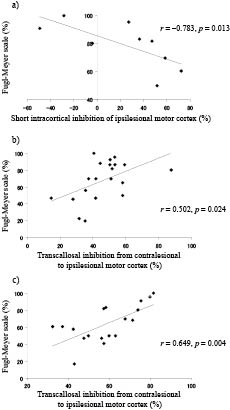
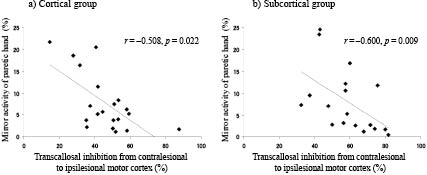
Table III shows the correlations between TMS parameters and motor function of the paretic hand. SICI of the ipsilesional motor cortex was negatively correlated with the FMS score of the paretic hand in the subcortical (Fig. 1a; r = –0.783, p = 0.013), but not the cortical group (r = –0.483, p = 0.187). TCI from the contralesional to the ipsilesional motor cortex was positively correlated with the FMS score of the paretic hand in both the cortical (Fig. 1b; r = 0.502, p = 0.024) and the subcortical groups (Fig 1c; r = 0.649, p = 0.004). There was a negative correlation between TCI to the ipsilesional motor cortex and mirror activity of the paretic hand in both the cortical (Fig. 2a; r = –0.508, p = 0.022) and the subcortical groups (Fig 2b; r = –0.600, p = 0.009). There was no significant correlation between TCI from the ipsilesional to the contralesional motor cortex and mirror activity of the non-paretic hand in either group.
| Table III. Correlations between transcranial magnetic stimulation parameters (TMS) and Fugl-Meyer scale (correlation coefficient and p-values) |
| TMS parameters | Fugl-Meyer scale |
| Ipsilesional hemisphere (stimulation site) | Contralesional hemisphere (stimulation site) |
| Cortical (n = 9) | Subcortical (n = 9) | Cortical (n = 20) | Subcortical (n = 18) |
| rMT | –0.497 (0.173) | –0.033 (0.933) | 0.038 (0.873) | 0.143 (0.570) |
| MEPs | 0.267 (0.488) | –0.183 (0.637) | –0.251 (0.285) | –0.060 (0.813) |
| SICI | –0.483 (0.187) | –0.783 (0.013)* | –0.121 (0.612) | –0.162 (0.521) |
| ICF | 0.300 (0.433) | 0.550 (0.125) | 0.403 (0.078) | 0.054 (0.832) |
| TCI | –0.200 (0.606) | –0.250 (0.516) | 0.502 (0.024)* | 0.649 (0.004)** |
| *p < 0.05; **p < 0.01. rMT: resting motor threshold; MEP: motor-evoked potentials; SICI: short intracortical inhibition; ICF: intracortical facilitation; TCI: transcallosal inhibition. |
Fig. 1. Correlation between inhibitory function and motor function. (a) There was a negative correlation between intracortical inhibition of the ipsilesional motor cortex and the Fugl-Meyer Scale score in the subcortical group. There was a significant positive correlation between transcallosal inhibition from the contralesional to the ipsilesional motor cortex and the Fugl-Meyer Scale score in both (b) the cortical and (c) the subcortical groups.
Fig. 2. The correlation between transcallosal inhibition and mirror activity of the paretic hand. There was a negative correlation between transcallosal inhibition from the contralesional to the ipsilesional motor cortex and mirror activity of the paretic hand in both (a) the cortical and (b) the subcortical groups.
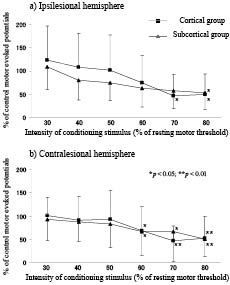
Fig. 3 shows the change in SICE in the cortical and the subcortical group. A repeated-measures ANOVA for SICE showed no significant interaction between INTENSITY and STIMULATION SITE (F (15, 260) = 0.884, p = 0.582) or STIMULATION SITE (F (3, 52) = 0.142, p = 0.935), but a significant effect of INTENSITY (F (5, 260) = 21.462, p < 0.001), reflecting that SICE had not been influenced by the stimulation site. Post-hoc analysis revealed that a strong conditioning stimulus could reduce SICE (Fig. 3).
Fig. 3. Short interval cortical excitability. The strong conditioning stimulus reduced the amplitude of motor evoked potentials (MEPs) in short interval cortical excitability in all groups. A significant reduction in the amplitude of the MEPs is indicated by asterisks. Error bar: standard deviation.
Discussion
This study revealed that the inhibitory function of the ipsilesional motor cortex correlated negatively with motor function of the paretic hand in subcortical stroke patients. The inhibition from the contralesional to the ipsilesional motor cortex correlated positively with motor function of the paretic hand; in contrast, the inhibition from the contralesional to the ipsilesional motor cortex correlated negatively with mirror activity of the paretic hand in both cortical and subcortical stroke patients.
Several studies have reported disinhibition of the ipsilesional motor cortex in the acute stage of both cortical and subcortical stroke (2, 10). However, whether the inhibitory function of the ipsilesional motor cortex normalizes or remains decreased in the chronic stage remains controversial (11, 12). The correlation between inhibitory function and motor function is also poorly understood. In this study, we have revealed that the inhibitory function of the ipsilesional motor cortex was correlated negatively with the motor function of the paretic hand in only subcortical stroke patients, but not cortical stroke patients, in the chronic stage. Considering these findings, the continuous disinhibition of the ipsilesional motor cortex in subcortical stroke patients may promote the best possible recovery of motor function by facilitating the plasticity of the non-damaged motor cortex in the ipsilesional hemisphere (4); in contrast, the inhibitory function of the ipsilesional motor cortex in cortical stroke patients may be influenced more by direct cortical damage than compensatory mechanisms in the chronic stage.
The problem with the SICI methods is that it was difficult to decide whether a reduced SICI indicated weak inhibitory or strong excitatory cortical function solely on the basis of the SICI paradigm. To avoid this problem, we used the SICE paradigm that could evaluate the inhibitory and excitatory circuits in more detail. The influence of the excitatory function has been shown to be superior to that of the inhibitory function at a strong conditioning stimulus in the SICE paradigm (7). If only the excitatory function increases and the inhibitory function remains unchanged, the amplitude of SICE is small at a weak conditioning stimulus and large at a strong conditioning stimulus (7). However, the amplitude of SICE was reduced according to the intensity of the conditioning stimulus in this study. Therefore, the reduction in SICI of the ipsilesional motor cortex implies the loss of inhibitory function and not an epiphenomenon caused by modified neuronal circuits shifting toward excitatory activity.
TCI from the contralesional to the ipsilesional motor cortex was more prominent in patients with greater motor function during movement. This finding is not consistent with that of previous study, which reported a negative correlation between TCI at pre-movement and the motor function of the paretic hand (5). These differences may have resulted from the differing methods and TCI mechanisms employed in our and previous study (13). A recent study reported that TCI could inhibit unwanted mirror activity during intended unimanual motor tasks (14). Consistent with this report, TCI to the ipsilesional motor cortex was correlated negatively with the mirror activity of the paretic hand in our study. Therefore, TCI to the ipsilesional motor cortex during movement may play a neurophysiological role in the inhibition of mirror movement of the paretic hand. To clarify this hypothesis, further studies are required to evaluate the change in mirror activity when TCI to the ipsilesional motor cortex is reduced by using inhibitory repetitive TMS over the contralesional motor cortex (14). We propose that TCI to the ipsilesional motor cortex may be important for mirror movement of the paretic hand; however, we agree with the hypothesis that TCI to the ipsilesional motor cortex may inhibit motor function in some stroke patients (5). Considering these findings, TCI to the ipsilesional motor cortex may be influenced by a balance between motor function and mirror movement in the paretic hand during the process of reorganization after stroke. That is to say, TCI to the ipsilesional motor cortex may be strong to inhibit mirror movement in patients with good motor function; in contrast, TCI in patients with poor motor function may be weak to improve motor function without inhibition of mirror movement.
The neurophysiological results of this study may help improve individualized rehabilitation strategies after stroke. Recent study has reported that inhibitory neuromodulation of the contralesional motor cortex could improve the motor function of the paretic hand by a reduction in TCI to the ipsilesional motor cortex (9). Therefore, inhibitory neuromodulation of the contralesional motor cortex may be especially effective for stroke patients with good motor function who had strong TCI, although the mirror activity of the paretic hand may increase. In addition, for subcortical stroke patients with disinhibition of the ipsilesional motor cortex, intense use of the paretic limb, such as constraint-induced movement therapy, may promote motor recovery by inducing use-dependent reorganization (15). In contrast, inhibitory neuromodulation of the contralesional motor cortex may be less effective in stroke patients with poor motor function, because these patients already have weak TCI before the neuromodulation interventions. The functional imaging study has reported that the contralesional motor cortex is engaged during paretic hand movements in stroke patients with poor motor function (16). Therefore, therapy aimed at increasing the excitability of the contralesional motor cortex may be effective for motor recovery of stroke patients with poor motor function. However, to our knowledge, there is no report that a neuromodulatory approach that increases the excitability in only the contralesional motor cortex can enhance motor recovery, ignoring the importance of the balance between bilateral hemispheres (17). If excitability is increased only in the contralesional motor cortex, the weak TCI to the ipsilesional motor cortex in stroke patients with poor motor function may become strong and inhibit the function of the ipsilesional motor cortex. Therefore, bilateral movement training that engages and balances both hemispheres may be effective for stroke patients with poor motor function (18).
Acknowledgements
We thank Mami Onodera for technical support. This work was supported by research project grant-in-aid for scientific research of No. 20700420 from the Japan Society for the Promotion of Science.
References